Ганглии нервной системы. Передача возбуждения на вегетативный ганглий
Вегетативные ганглии представляют собой скопление многочислен-ных мультиполярных нервных клеток.
Величина вегетативных ганглиев существенно варьирует. В связи с этим различают крупные, средней величины, мелкие и очень мелкие (микроганглии) ганглии.
Необходимо отметить, что кроме анатомически обособленных ганглиев, по ходу вегетативных ветвей периферических нервов встречается большое количество нервных клеток, подобных нервным клеткам вегетативного ганглия. Эти нейроны, мигрирующие сюда в ходе эмбриогенеза, локализуются по ходу нервов поодиночке или образуют небольшие группы – микроганглии.
Вегетативный ганглий с поверхности покрыт фиброзной соединительнотканной капсулой, от которой внутрь отходят многочисленные прослойки соединительной ткани, образующей строму узла. По этим прослойкам в узел проходят кровеносные сосуды, питающие его и образующие в нем капиллярную сеть. В капсуле и строме узла часто вблизи кровеносных сосудов встречаются рецепторы – диффузные, кустиковидные или инкапсулированные.
Мультиполярные нервные клетки вегетативного ганглия впервые были описаны А.С. Догелем. При этом Догель выделил 3 типа нервных клеток вегетативного ганглия, которые получили названияклеток Догеля I , II, III типа . Морфофункциональные характеристики клеток Догеля существенно разнятся.
Клетки Догеля I типа по функциональному значению являются эффекторными (двигательными) нейронами. Это более или менее крупные нервные клетки, с несколько короткими дендритами, не выходящими за пределы данного ганглия. Аксон этих клеток более длинный выходит за пределы ганглия и направляется к рабочему аппарату – гладкомышечным клеткам, железистым клеткам, образуя на них двигательные (или соответственно секреторные) нервные окончания. Аксоны и дендриты клеток ДогеляIтипа являются безмякотными. Дендриты часто образуют пластинчатые расширения, на которых (как и на теле клетки) располагаются синаптические окончания, образующиеся разветвлениями преганглионарного нервного волокна.
Тела нейронов в вегетативном ганглии, в отличие от спиномозгового ганглия, располагаются беспорядочно по всему узлу и более рыхло (т.е. более редко). На препаратах, окрашенных гематоксилином или другими общегистологическими красителями, отростки нервных клеток остаются не выявленными, а клетки имеют такую же округлую безотросчатую форму, как в спиномозговых узлах. Тело каждой нервной клетки (как и в спинальном ганглии) окружено слоем уплощенных элементов олигодендроглии – слоем сателлитов.
К наруже от слоя сателлитов имеется еще тонкая соединительно-тканная капсула. Клетки Догеля Iтипа являются основной клеточной формой вегетативных ганглиев.
Клетки Догеля II типа – это также мультиполярные нервные клетки, с несколькими длинными дендритами и нейритом, уходящим за пределы данного ганглия в соседние ганглии. Аксон с поверхности покрыт миелином. Дендриты этих клеток начинаются рецепторными аппаратами в гладких мышцах. С функциональной точки зрения клетки ДогеляIIтипа являются чувствительными. В отличие от чувствительных псевдоуниполярных нервных клеток спиномозгового узла клетки ДогеляIIтипа, по-видимому, образуют рецепторное (афферентное) звено местных рефлекторных дуг, замыкаемых без захода нервного импульса в центральную нервную систему.
Клетки Догеля III типа представляют собой местные ассоциативные (вставочные) элементы, соединяющие своими отростками несколько клетокIиIIтипа. Их дендриты короткие, но более длинные, чем у клетокIтипа, не выходят за пределы данного ганглия, а образуют корзинчатые разветвления, оплетающие тела других клеток данного ганглия. Нейрит клетки ДогеляIIIтипа идет в другой ганглий и там вступает в синаптическую связь с клеткамиIтипа. Следовательно, клеткиIIIтипа входят в качестве ассоциативного звена в местные рефлекторные дуги.
Нельзя не отметить, существует такая точка зрения, что клетки Догеля IIIтипа имеют рецепторную или эффекторную природу.
Соотношение численности клеток Iи II типов Догеля в различных вегетитивных ганглиях неодинаково. Парасимпатические ганглии, в отличие от симпатических ганглиев, характеризуются преобладанием клеток с короткими внутрикапсулярными дендритами, отсутствием или малым количеством пигмента в клетках. Кроме того в парасимпатических ганглиях, как правило, тела лежат значительно компактнее, чем в симпатических ганглиях. Кроме того, в симпатических ганглиях имеютсяМИФ-клетки (мелкие клетки с интенсивной флюоросценцией).
Через вегетативный ганглий проходят проводящие пути трех видов: центростремительные, центробежные и периферические (местные) рефлекторные.
Центростремительные пути образованы чувствительными отростками псевдоуниполярных клеток спинального ганглия, начинающихся рецепторами в иннервируемых тканях, а также внутри ганглия. Эти волокна проходят транзитом через вегетативные ганглии.
Центробежные пути представлены преганглионарными волокнами, которые многократно ветвятся в вегетативном узле и образуют синапсы на многих клеточных телах эффекторных нейронов. Например, в верхнем шейном узле соотношение числа преганглионарных волокон, вступивших в него, к постганглионарным равно 1:32. Это явление приводит, при возбуждении преганглионарных волокон, к резкому расширению области возбуждения (геперализация эффектора). Благодаря этому, сравнительно небольшое количество центральных вегетативных нейронов обеспечивает нервными импульсами все органы и ткани. Итак, например, при раздражении у животного преганглионарных симпатических волокон, проходящих через передние корешки IYгрудного сегмента, может наблюдаться сужение сосудов кожи головы, шеи, расширение коронарных сосудов, сужение сосудов кожных покровов передней конечности, сосудов почки и селезенки.
Продолжение этих путей составляют постганглионарные волокна, достигающие иннервируемых тканей.
Периферические (местные) рефлекторные пути начинаются в тканях разветвлениями отростков собственных чувствительных нейронов вегетативных ганглиев (т.е. клетками IIтипа Догеля). Нейриты же этих кле-ток заканчиваются на клетках ДогеляIтипа, чьи постганглионарные волокна входят в состав центробежных путей.
Морфологическим субстратом рефлекторной деятельности вегетативной нервной системы является рефлекторная дуга. Для рефлектор-ной дуги вегетативной нервной системы характерны все три звена – рецепторное (афферентное), вегетативное (ассоциативное) и эффекторное (двигательное), но локализация их иная чем в соматической.
Интересно отметить, что многие морфологи и физиологи указывают как на отличительный признак вегетативной нервной системы, отсутствие в ее составе собственного афферентного (рецепторного) звена, т.е. они считают, что чувствительная иннервация внутренних органов, сосудов и т.д. осуществляется дендритами псевдоуниполярных клеток спинального ганглия, т.е. соматической нервной системы.
Правильнее считать, что спиномозговые узлы содержат нейроны, иннервирующие скелетную мускулатуру, кожу (т.е. нейроны соматической нервной системы), так и нейроны, иннервирующие все внутренние органы, сосуды (т.е. вегетативные нейроны).
Одним словом, аффекторное звено, как и в соматической (анимальной) нервной системе, в вегетативной нервной системе представлено клеткой, лежащей в спиномозговом узле.
Тело нейрона ассоциативного звена располагается, в отличие от соматической рефлекторной нервной дуги, не в области заднего рога, а в боковых рогах серого вещества, и аксон этих клеток выходит за пределы мозга и оканчивается в одном из вегетативных ганглиев.
Наконец, наибольшие отличия между анимальной и вегетативной рефлекторными дугами наблюдаются в эфферентном звене. Так, тело эфферентного нейрона в соматической нервной системе находится в сером веществе спинального или головного ганглия и лишь его аксон идет на периферию в составе того или иного черепно или спиномозгового нерва. В вегетативной системе тела эффекторных нейронов находятся на периферии: они либо разбросаны по ходу некоторых нервов, либо образуют скопления – вегетативные ганглии.
Таким образом, для вегетативной нервной системы, в силу такой локализации эффекторных нейронов, характерно наличие, по крайней мере, одного перерыва эфферентного пути, который проходит в вегетативном ганглии, т.е. здесь нейриты вставочных нейронов, контактируют с нейронами эффекторными, образуя на их телах и дендриты синапсы. Следовательно, вегетативные ганглии представляют собой периферические нервные центры. Этим они принципиально отличаются от спинальных ганглиев, которые не являются нервными центрами, т.к. в них нет синапсов и не происходит переключения нервных импульсов.
Таким образом, спиномозговые узлы являются смешанными образованиями, анимально-вегетативными.
Особенностью рефлекторной дуги симпатической нервной системы является наличие коротких преганглионарных волокон и очень длинных постганглионарных волокон.
Особенностью рефлекторной дуги парасимпатической нервной системы является, напротив, наличие очень длинных преганглионарных и очень коротких постганглионарных волокон.
Основные функциональные различия симпатической и парасимпатической систем заключаются в следующем. Медиатором, т.е. веществом, образующимся в области синапсов и осуществляющим химическую передачу импульса, в симпатических нервных окончаниях является симпатин (вещество, тождественное гормону мозгового вещества надпочечника – ноадреналину).
Медиатором в парасимпатических нервных окончаниях является «вещество вагуса» (вещество, тождественное ацетилхолину). Впрочем эта разница касается только постганглионарных волокон. Синапсы, образованные преганглионарными волокнами и в симпатической и парасимпатической системах холинергичны, т.е. в качестве медиатора они образуют холиноподобное вещество.
Названные химические вещества – медиаторы и сами по себе, даже без раздражения вегетативных нервных волокон, вызывают в рабочих органах эффекты, аналогичные действию соответствующих вегетативных нервных волокон. Так, ноадреналин при введении в кровь ускоряет сердцебиение, но замедляет перистальтику кишечного тракта, а ацетилхолин – наоборот. Ноадреналин вызывает сужение, а ацетилхолин – расширение просвета сосудов.
Холинергичны и синапсы, образуемые волокнами соматической нервной системы.
Деятельность вегетативной нервной системы находится под контролем коры больших полушарий, а также подкорковых вегетативных центров полосатого тела и, наконец, вегетативных центров промежуточного мозга (ядро гипоталамуса).
В заключении необходимо отметить, что учение о вегетативной нервной системе большой вклад внесли и советские ученые Б.И. Лаврентьев, А.А. Заварзин, Д.И. Голуб, удостоенные государственных премий.
Литература:
Жаботинский Ю.М. Нормальная и патологическая морфология вегетативных ганглиев. М.,1953
Заварзин А.А. Очерк по эволюционной гистологии нервной системы. М-Л,1941
А.Г. Кнорре, И.Д.Лев. Вегетативная нервная система. Л.,1977,с.120
Колосов Н.Г. Иннервация пищеварительного тракта человека. М-Л,1962
Колосов Н.Г. Вегетативный узел. Л.,1972
Колосов Н.Г., Хабарова А.Л. Структурная организация вегетативных ганглиев. Л.,Наука, 1978.-72с.
Кочетков А.Г., Кузнецов Б.Г., Коновалова Н.В. Вегетативная нервная система. Н-Новгород, 1993.-92с.
Мельман Е.П. Функциональная морфология иннервация органов пищеварения. М.,1970
Ярыгин Н.Е. и Ярыгин В.Н. Патологические и приспособительные изменения нейрона. М.,1973.
Вегетативная нервная система (ВНС) в основном обеспечивает иннервацию внутренних органов.
Делится на:
Симпатический отдел
Парасимпатический отдел
Метасимпатический (Энтеральный)
Отличия вегетативной нервной системы от соматической НС:
- Не находится под контролем сознания
- Возможность автономного функционирования (даже при полном нарушении связи с ЦНС)
- Генерализованный характер распространения возбуждения в периферическом отделе ВНС (особенно в симпатическом отделе).
- Наличие вегетативного ганглия в эфферентном отделе рефлекторной дуге. Таким образом, эфферентная часть ВНС представлена двумя нейронами: преганглионарный нейрон в пределах ЦНС (ствол мозга, спинной мозг), постганглионарный нейрон в вегетативном ганглии. Т.е. тела последних нейронов вегетативных дуг вынесены за пределы ЦНС.
- Низкая скорость проведения нервного импульса (преганглионарные волокна типа В, постганглионарные типа С)
- Ткани-мишени для ВНС: гладкомышечные клетки, поперенополосатая сердечная мышца, железистая ткань (для соматической — поперечнополосатая скелетная МТ). Симпатические волокна способны влиять на гликогенолиз в печени и липолиз в жировых клетках (метаболический эффект)
Обычно внутренние органы имеют двойную иннервацию: симпатическую и парасимпатическую, однако мочевой пузырь и ресничная мышца получают в основном парасимпатическую, сосуды, потовые железы, волоковые мышцы кожи, селезёнка, матка, мозг, органы чувств, надпочечники – только симпатическую.
Высшие вегетативные центры
структуры лимбической системы, базальные ганглии, КГМ, гипоталамус (передние ядра – зона парасимпатических ядер, задние — зона симпатических ядер), центральное серое вещество среднего мозга, ретикулярная формация (её нейроны формируют жизненно важные центры продолговатого мозга ССЦ, ДЦ).
Нервные центры (центральный отдел) симпатической нервной системы – промежуточно-латеральные ядра боковых рогов спинного мозга C VIII — L II — III
Нервные центры (центральный отдел) парасимпатической нервной системы – вегетативные ядра III пары (глазодвигательный нерв – Ядро Якубовича), VII (лицевой нерв – верхнее слюноотделительное), IX (языкоглоточный нерв – нижнее слюноотделительное), X (блуждающий нерв – заднее ядро), промежуточно-латеральные ядра спинного мозга S II -S IV
На уровне рабочих отделов заложены эфферентные клетки, аксоны которых идут не сразу к рабочему органу, в отличие от соматических, а прерываются в периферическом вегетативном ганглии. Здесь они переключается на последние нейроны, Волокна нейронов спинного мозга называются преганглионарными. Преганглионарные волокна переключаются в вегетативном ганглии на следующий нейрон, аксон которого носит название постганглионарного.
Симпатический вегетативный ганглий
Ганглий покрыт сверху капсулой. Здесь имеются следующие клетки:
- Чувствительные нейроны
- Эфферентные нейроны
- Хромаффинные клетки, выделяющие катехоламины (регулируют уровень возбудимости клеток узла.
Функции ганглия: проводниковая, замыкательная и рецепторная.
Нейроны вегетативного ганглия обладают теми же свойствами, что и нейроны центральной нервной системы.
Парасимпатический вегетативный ганглий
Ганглий покрыт сверху капсулой. В нём имеются следующие клетки:
- Чувствительные — клетки Догеля 2-го типа, их рецепторы могут быть механо-, термо-, хемочувствительными.
- Эффекторные нейроны – клетки Догеля 1-го типа, имеют много коротких дендритов и один аксон, уходящий за пределы ганглия.
- Вставочные – клетки Догеля 3-го типа.
- В ганглии имеются также хромаффинные клетки, выделяющие катехоламины, возможно, серотонин, АТФ, нейропептиды (регуляторная функция).
Физиология вегетативного ганглия
(переключение с преганглионарных волокон на постганглионарные)
- Низкая лабильность нейронов вегетативных ганглиев (10-15 импульсов в секунду), у соматических 200 имп/сек.
- Длительная синаптическая задержка, больше в 5 раз.
- Большая длительность ВПСП (20-50 мс), длительность потенциала действия 1,5-3 мс из-за продолжительной следовой гиперполяризации нейронов ганглия.
- Большую роль имеет пространственная и последовательная суммация.
- Медиатор: в вегетативных ганглиях – преганглионарные нейроны выделяют АХ.
- На уровне ганглия хорошо развита конвергенция и дивергенция (мультипликация).
Симпатический отдел вегетативной нервной системы
Симпатические вегетативные ганглии расположены в симпатическом стволе, превертебральных узлах, узлах сплетений (брюшное аортальное, верхнее и нижнее подчревное).
Преганглионарные волокна короткие, сильно ветвятся. Постганглионарные волокна длинные тонкие многократно ветвятся, образуют сплетения. Хорошо развита мультипликация.
Медиатор постганглионарных адренергических симпатических волокон – НА (90%), адреналин (7%), дофамин (3%). Медиатор стойкий, длительно проявляет свою активность. НА связывается с α и β-адренорецепторами органов-эффекторов. Классификация основана на чувствительности их к фарм.препаратам: α-адренорецепторы блокируются фентоламином, β — пропранололом. Адренорецепторы имеются не только на органах, иннервируемых симпатическими волокнами (сердце, жировая ткань, сосуды, мышца-дилататор зрачка, матка, семявыносящий проток, кишечник) (α 1 и β 1), но и вне синапсов (на тромбоцитах, скелетные мышцы, эндокринные и экзокринные железы) (α 2 и β 2), а также на пресинаптческой мембране.
Передача возбуждения осуществляется быстрее, чем по симпатическому отделу. Влияния краткосрочные.
Влияния:
- Постоянное (тоническое)
- Фазное (пусковое) – резкое изменение функции (зрачковый рефлекс)
- Адаптационно-трофическое
Адапционно-трофическое влияние симпатической нервной системы Орбели-Гинецинского
Это приспособление обменных процессов к уровню функциональной активности. Идею о трофическом влиянии сформулировал И.П.Павлов. В опыте на собаке обнаружил симпатическую веточку, идущую к сердцу, раздражение которой вызывало усиление сердечных сокращений, без изменения частоты. Усиление сокращений утомлённой мышцы связано с активацией обменных (трофических) процессов под влиянием НА. Он активирует специфические рецепторы мембраны мышечных волокон, запускает каскад химических реакции в цитоплазме, ускоряющих синтез макроэргов, повышает возбудимость периферических рецепторов. Предполагается наличие трофогенов в нервных окончаниях. К трофогенам относятся нуклеотиды, некоторые аминокислоты, простагландины, катехоламины, серотонин, АХ, сложные липиды, ганглиозиды.
Парасимпатическая отдел вегетативной нервной системы
Парасимпатические вегетативные ганглии (далеко от ЦНС) расположены внутри органов (интрамурально) или околоорганно (ресничный, крылонёбный, ушной, подъязычный, поднижнечелюстной узлы), в узлах сплетений.
Преганглионарные волокна длинные, слабо ветвятся. Постганглионарные волокна короткие, мало ветвятся. Мультипликация развита слабо.
Медиатор постганглионарных парасимпатических волокон АХ.
Ацетилхолин на клетках-эффекторах связывается М-холинорецепторами. М-холинорецепторы возбуждаются мускарином, блокируются ядом кураре.
Ацетилхолин – нестойкий медиатор, основная часть разрушается ацетилхолинэстеразой до холина и ацетата, которые затем захватываются пресинаптической мембраной и используются для синтеза. Меньшая часть диффундирует в интерстиций и кровь.
Влияния:
- Постоянное (тоническое)
- Фазное (пусковое) – резкое изменение функции (торможение работы сердца, активация перистальтики, сужение зрачка)
Тонус вегетативных центров
Многие преганглионарные и ганглионарные нейроны обладают постоянной активностью, называемой тонусом. В покое частота электрических импульсов в вегетативных волокнах 0,1-5 имп/с. Тонус вегетативных нейронов подвержен суточным колебаниям: симпатотонус днём выше, ночью ниже, во время сна повышается тонус парасимпатических волокон. Симпатотонус обеспечивает постоянный тонус сосудов. Тоническое влияние блуждающего нерва (ваготонус) на сердце постоянно сдерживает ЧСС. Чем выше двигательная активность у человека, тем больше выражен парасимпатический тонус (уменьшение ЧСС у спортсменов). Причины вегетативного тонуса:
- Спонтанная активность. Высокий уровень спонтанной активности характерен для нейронов РФ.
- Поток афферентных импульсов от различных рефлексогенных зон.
- Действие БАВ и метаболитов
Вегетативные рефлексы. Классификация:
По уровню замыкания:
- центральные (соматовегетативный рефлекс- имеет общую афферентную часть с соматическим рефлексом)
- периферические, автономные (дуга рефлекса может замыкаться вне ЦНС в вегетативном ганглии интраорганно или экстраорганно, возможно существование аксон-рефлекса)
По расположению рецепторов:
Интероцептивные (механо-, хемо-, термо-, ноце-, полимодальные рецепторы)
а) Висцеро-висцеральные (каротидный синус, солнечное сплетение, перистальтика)
б) Висцеро-кутанные (соответственно зонам Захарьина-Геда)
в) Висцеро-моторные (раздражение интерорецепторов может вызывать моторные реакции).
-
Автономные (вегетативные) нервные узлы (ганглии) могут располагаться вдоль позвоночника (паравертебральные ганглии) или впереди него (превертебральные ганглии), а также в стенке органов: сердца, бронхов, пищеварительного тракта, мочевого пузыря и других (интрамуральные ганглии) или вблизи их поверхности. Иногда они имеют вид мелких (от нескольких клеток до нескольких десятков клеток) скоплений нейронов, расположенных по ходу некоторых нервов или лежащих интрамурально (микроганглии). К вегетативным узлам подходят преганглионарные волокна (миелиновые), содержащие отростки клеток, тела которых лежат в центральной нервной системе. Эти волокна сильно ветвятся и образуют многочисленные синаптические окончания на клетках вегетативных узлов. Благодаря этому осуществляется конвергенция большого числа терминалей преганглионарных волокон на каждый нейрон ганглия. В связи с наличием синаптической передачи вегетативные узлы относят к нервным центрам ядерного типа.
Вегетативные нервные узлы по функциональному признаку и локализации разделяются на:
симпатические;
парасимпатические.
Симпатические нервные узлы (пара- и превертебральные) получают преганглионарные волокна от клеток, расположенных в вегетативных ядрах грудных и поясничных сегментов спинного мозга. Нейромедиатором преганглионарных волокон является ацетилхолин, а постганглионарных — норадреналин (за исключением потовых желез и некоторых кровеносных сосудов, имеющих холинергическую симпатическую иннервацию). Помимо этих нейромедиаторов, в узлах выявляются энкефалины, вещество Р, соматостатин, холецистокинин.
Парасимпатические нервные узлы (интрамуральные, лежащие вблизи органов или узлы головы) получают преганглионарные волокна от клеток, расположенных в вегетативных ядрах продолговатого и среднего мозга, а также крестцового отдела спинного мозга. Эти волокна покидают центральную нервную систему в составе 3, 7, 9, 10 пар черепномозговых нервов и передних корешков крестцовых сегментов спинного мозга. Нейромедиатором пре- и постганглионарных волокон является ацетилхолин. Кроме него роль медиаторов в этих ганглиях играют серотонин, АТФ, возможно, некоторые пептиды.
Большинство внутренних органов имеет двойную вегетативную иннервацию, то есть получают постганглионарные волокна от клеток, расположенных как в симпатических, так и в парасимпатических узлах. Реакции, опосредуемые клетками симпатических и парасимпатических узлов, часто имеют противоположную направленность, например: симпатическая стимуляция усиливает, а парасимпатическая — тормозит сердечную деятельность.
Общий план строения симпатических и парасимпатических нервных узлов сходен. Вегетативный узел покрыт соединительнотканной капсулой и содержит диффузно или группами расположенные тела мультиполярных нейронов, их отростки в виде безмиелиновых или, реже, миелиновых волокон и эндоневрий. Тела нейронов имеют неправильную форму, содержат эксцентрично расположенное ядро, окружены (обычно не полностью) оболочками из глиальных клеток-сателлитов (мантийных глиоцитов). Часто встречаются многоядерные и полиплоидные нейроны.
Интрамуральные узлы и связанные с ними проводящие пути в виду их высокой автономии, сложности организации и особенностей медиаторного обмена некоторыми авторами выделяются в самостоятельный метасимпатический отдел вегетативной нервной системы. В частности, общее число нейронов в интрамуральных узлах кишки выше, чем в спинном мозге, а по сложности их взаимодействия в регуляции перистальтики и секреции их сравнивают с миникомпьютером.
В интрамуральных узлах описаны нейроны трех типов:
длинноаксонные эфферентные нейроны (клетки Догеля I типа) численно преобладают. Это крупные или средних размеров эфферентные нейроны с короткими дендритами и длинным аксоном, направляющимся за пределы к рабочему органу, на клетках которого он образует двигательные или секреторные окончания;
равноотростчатые афферентные нейроны (клетки Догеля II типа) содержат длинные дендриты и аксон, уходящий за пределы данного ганглия в соседние и образующий синапсы на клетках I и III типов. Эти клетки, по-видимому, входят в качестве рецепторного звена в состав местных рефлекторных дуг, которые замыкаются без захода нервного импульса в центральную нервную систему. Наличие таких дуг подтверждается сохранением функционально активных афферентных, ассоциативных и эфферентных нейронов в трансплантированных органах (например, сердце);
ассоциативные клетки (клетки Догеля III типа) — местные вставочные нейроны, соединяющие своими отростками несколько клеток I и II типа, морфологически сходные с клетками Догеля II типа. Дендриты этих клеток не выходят за пределы узла, а аксоны направляются в другие узлы, образуя синапсы на клетках I типа.
Подробности
Ганглии представляют собой скопления мультиполярных (один аксон и несколько дендритов) нейронов (от нескольких клеток до десятков тысяч). Экстраорганные (симпатические) ганглии имеют хорошо выраженную соединительнотканную капсулу, как продолжение периневрия. Парасимпатические ганглии находятся, как правило, в интрамуральных нервных сплетениях. Ганглии интрамуральных сплетений, как и другие вегетативные узлы, содержат вегетативные нейроны местных рефлекторных дуг. Мультиполярные нейроны диаметром 20-35 мкм расположены диффузно, каждый нейрон окружен глиоцитами ганглия.
Кроме того, описаны нейроэндокринные, хеморецепторные, биполярные, а у некоторых позвоночных и униполярные нейроны . В симпатических ганглиях имеются мелкие интенсивно флюоресцирующие клетки (МИФ-клетки) с короткими отростками и большим количеством гранулярных пузырьков в цитоплазме. Они выделяют катехоламины и оказывают тормозящее влияние на передачу импульсов с преганглионарных нервных волокон на эфферентный симпатический нейрон. Эти клетки называют интернейронами.
Среди крупных мультиполярных нейронов вегетативных ганглиев различают: двигательные (клетки Догеля 1-го типа), чувствительные (клетки Догеля П-го типа) и ассоциативные (клетки Догеля Ш-го типа) . Двигательные нейроны имеют короткие дендриты с пластинчатыми расширениями ("рецептивные площадки"). Аксон этих клеток очень длинный, уходит за пределы ганглия в составе постганглионарных тонких безмиелиновых нервных волокон и оканчивается на гладких миоцитах внутренних органов. Клетки 1-го типа называют длинноаксонными нейронами. Нейроны П-го типа - равноотростчатые нервные клетки. От их тела отходят 2-4 отростка, среди которых различить аксон трудно. Не разветвляясь, отростки уходят далеко от тела нейрона. Их дендриты имеют чувствительные нервные окончания, а аксон оканчивается на телах двигательных нейронов в соседних ганглиях. Клетки П-го типа являются чувствительными нейронами местных вегетативных рефлекторных дуг. Клетки Догеля Ш-го типа по форме тела похожи на вегетативные нейроны П-го типа, но их дендриты не выходят за пределы ганглия, а нейрит направляется в другие ганглии. Многие исследователи считают эти клетки разновидностями чувствительных нейронов.
Таким образом, в периферических вегетативных ганглиях имеются местные рефлекторные дуги, состоящие из чувствительных, двигательных и, возможно, ассоциативных вегетативных нейронов.
Интрамуральные вегетативные ганглии в стенке пищеварительного тракта отличаются тем, что в их составе, кроме двигательных холинергических нейронов, имеются тормозные нейроны. Они представлены адренергическими и пуринергическими нервными клетками. В последних медиатором является пуриновый нуклеотид. В интрамуральных вегетативных ганглиях встречаются также пептидергические нейроны, выделяющие вазоинтестинальный пептид, соматостатин и ряд других пептидов, с помощью которых осуществляются нейроэндокринная регуляция и модуляция деятельности тканей и органов пищеварительной системы.Ацетилхолин - никотиновые (блок кураре, гексаметоний), мускариновые (блок атропин) рецепторы. Активация рецепторов→генерирование ВПСП. Быстрый ВПСП (N-холиноцер)→открытие ионных каналов. Медленный ВПСП (М-холинорец)→подавление М-тока, обусловленного повышением К-проводимости.
Нейропептиды – действуют как нейромодуляторы.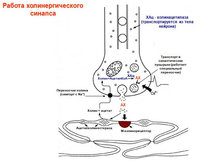
Энкефалины , вещество Р, люлиберин, нейротензин, соматостатин – симп. ганглии (+Ach)
Катехоламины (NA, дофамин) – нейромедиаторы мелких клеток с интенсивной флуоресценцией.
Нейропептид Y, соматостатин – симп. постганглионары.Симпатические постганглионары: NA, АТФ, нейропептид У.
α1→инозотолтрифосфат, диацилглицерол. α2→активация G-белка, ↓цАМФ.
β→G-белок→AC→цАМФ
Исключения : медиатор Ach, рецепторы мускариновые.
Парасимп. постганглионары: Ach, ВИП, NO, соматостатин, АТФ, опиоидные пептиды.
М1 (высокое сродство к пирензепину)-повышение секреции кислоты клетками желез желудка, М2 (низкое)- замедляют серд. ритм, секреция слезных и слюнные желез.
Разнообразное действие:
-Специфические втор. посредники: М2 может акт-ть IP3, а может инг-ть AC, уменьшая цАМФ.
-Действие на К и Са-каналы
-На эндотелии образуется NO→гуанилатциклаза→цГМФ→ цГМФ-зависимая протеинкиназа→расслабление глажких мышц.Вегетативная нервная система поддерживает гомеостаз. ВНС управляет такими висцеральными функциями, как кровообращение, пищеварение и выделение, главным образом без условного или сознательного контроля. ВНС также модулирует функцию эндокринных желез, регулирующих метаболизм. ВНС имеет сенсорные и моторные компоненты и разделяется на симпатическую и парасимпатическую системы. Первые нейроны симпатической системы расположены в промежуточных рогах тораколюмбального отдела спинного мозга; синапс со вторым набором нейронов находится в параили превертебральном симпатическом ганглии. В парасимпатической системе первые нейроны расположены либо в черепно-мозговом нерве, в автономных ядрах, либо в промежуточном роге сакрального отдела спинного мозга; синапс со вторым набором нейронов находится или в автономном ганглии (в случае черепно-мозговых нервов), или в эффекторной ткани непосредственно. ВНС имеет три главных компонента:
афферентный (центростремительный, чувствительный);
центральный объединяющий;
эфферентный.Афферентный компонент несет информацию от нейрональных физиологических рецепторов, расположенных в концах центростремительных нервов, к спинному мозгу и более высоким областям ЦНС. Большая часть этой информации обрабатывается в пределах гипоталамуса и других нижележащих областей мозга. После обработки соответствующий сигнал посылается от ЦНС вниз по эфферентным нервам к исполнительным органам (см. рис. 8.1, 8.9), названным так потому, что они отвечают на деятельность в ЦНС.
На основе различий анатомии и медиаторов эфферентную часть ВНС подразделяют на три системы:
парасимпатическую (холинергическую);
симпатическую (адренергическую);
неадренергическую нехолинергическую (НАНХ).Ацетилхолин - нейромедиатор холинергической системы. - нейромедиатор, высвобождаемый из пресинаптического окончания в автономном ганглии и в окончаниях нервов в исполнительном органе. Рецепторами для ацетилхолина служат холинорецепторы, которые подразделяют на мускариновые и никотиновые.
Норадреналин - нейромедиатор адренергической системы. Другая важная составляющая ВНС - адренергическая система. До сих пор неизвестно, какой нейромедиатор использовался в этой системе первоначально - эпинефрин или норэпинефрин. Сейчас известно, что за исключением надпочечников, которые секретируют эпинефрин (адреналин), нейроме-диатором в адренергической системе является норэпинефрин.
Ацетилхолин - ганглионарный медиатор для холинергической и адренергической систем. Эфферентные нервы и для холинергической, и для адренергической систем происходят из соответствующих частей ствола мозга и спинного мозга. Эфферентные нервы образуют синапс в ганглии, расположенном вне органа, где основным нейромедиатором является АХ:
В адренергической системе ганглии находятся в цепочке вблизи спинного мозга, известной как паравертебральная симпатическая цепочка;
в холинергической системе ганглий обычно располагается внутри или вблизи эффекторного органа.Несмотря на явное анатомическое различие , оба типа ганглиев используют АХ как основной ганглионарный нейромедиатор, активирующий никотиновые рецепторы.
Нейромедиаторы могут модулировать собственное высвобождение. Нейромедиаторы могут модулировать собственное высвобождение. Нейромедиаторы могут активировать пресинаптические рецепторы на нейроне, что ингибирует высвобождение самих нейромедиаторов.Учебное видео анатомии вегетативной нервной системы (ВНС)
При проблемах с просмотром скачайте видео со страницы- Вернуться в оглавление раздела " "